The following account is from Sarnat et al. (2015) and figure numbers refer to that publication.
Pheidole fervens is a medium to large sized species with long limbs. It belongs to the P. fervens clade along with its Australasian congeners P. cariniceps, P. hospes, P. impressiceps, and P. oceanica (Economo et al. 2015). The major workers have strong cephalic rugulae that become reticulated towards the posterior of the head and the minor workers have completely glossy heads with very long antennal scapes. Majors and minors of the species can be separated from those of P. megacephala and P. noda by the postpetiole which is not swollen compared to the petiole (Fig. 3), and the promesonotum which has the large anterior dome in addition to a distinct prominence on the posterior slope (Fig. 5, major; Fig. 43, minor). The minors of P. fervens can also be separated from those of P. megacephala by their larger size and longer antennal scapes (Fig. 39). The majors are easily distinguished from P. megacephala by the very sculptured head (Fig. 13).
Among species treated here, P. fervens is most easily confused with its close relative, P. indica, and the characters used to separate these two are subtle. For both subcastes, the promesonotal prominence is flatter in P. fervens (Fig. 49a, minor; Fig. 63a, major) compared to that of P. indica (Fig. 50a, minor; Fig. 64a, major). The eyes of P. fervens minors (Fig. 65) are relatively smaller than those of P. indica minors (Fig. 66), especially in comparison to antennal segment 10. The propodeal spines of P. fervens are weaker, narrower, and more downcurved in majors of P. fervens (Fig. 63b) compared to those of P. indica (Fig. 64b). Readers are referred to Eguchi (2004b; 2008) for characters used to separate P. fervens and P. indica from their Asian congeners.
In the Pacific Island region P. fervens is often confused with the nearly identical P. oceanica, which is native to that region. The carinae between eye and mandible are branching and reticulated in the majors of P. fervens (Fig. 67), versus parallel and not reticulated in those of P. oceanica (Fig. 68). This character was erroneously reversed in the key provided in Sarnat and Economo (2012). The minors are more difficult to separate, but in P. fervens the length of propodeal spine is equal to or less than the diameter of propodeal spiracle (Fig. 69), whereas in P. oceanica it is greater (Fig. 70).
For such a ubiquitous species across its native and introduced range, very little is known about the biology of Pheidole fervens. It is a very synanthropic species with a high tolerance for disturbance (Eguchi 2004b; Fischer and Fisher 2013; Martínez 1996), but can also thrive under some degree of canopy cover (Morrison 1996; Sarnat and Economo 2012). In Fiji, where it is likely a recent colonizer, it was collected most frequently in human dominated landscapes between 0–800 m, although several collections were also made from primary forest at low elevations. In Hawaii, where it is definitely an introduced species, it is more abundant locally in wet regions than P. megacephala (Gruner et al. 2003) and occurs in the hot lowlands only below 900 m (Reimer 1994). In the Philippines, P. fervens is found in irrigated lowlands (rice fields) where it is characterized as dominant species capable of displacing S. geminata in the dry season (Way et al. 1998). In Japan it occurs in open land grading to forest edge (Harada et al. 2009; Ogata 1981). Pheidole fervens recruits in large numbers to bait and forages both on the ground and on vegetation (Sarnat and Economo 2012). Baiting experiments on Pacific Islands found that P. fervens can act as a numerically and behaviorally dominant species capable of excluding other invasive ant species (including Anoplolepis gracilipes, Nylanderia bourbonica, and Tetramorium bicarinatum) from baits (Morrison 1996). Although foragers can be slow to discover food resources, once found they can recruit in large numbers and displace competing species (Morrison 1996). Experiments in China suggest that P. fervens can provide some degree of biotic resistance to the Red Imported Fire Ant (Solenopsis invicta) by acting in groups to dismember the limbs of individual fire ants (Chen et al. 2011). Martínez (1996) suggested the California population of P. fervens was polydomous, and Passera (1994) suggested the Hawaii population is unicolonial and polygynous, but detailed colony-level studies of the species are required to verify these claims. Wittenborn and Jeschke (2011) attributed their assertion that P. fervens practices dependent colony founding to Harris et al. (2005a), but we were unable to find any reference to colony foundation in that report and cannot substantiate their evidence.
Pheidole fervens can be a dominant species where it is locally abundant. Although few studies have measured the effect of P. fervens on native ecosystems, we predict that it could negatively impact native arthropods. We were unable to find documentation on the effect of P. fervens on agricultural systems, but it can be among the most abundant ant species in irrigated lowland crop systems such as rice fields. Pheidole fervens can also be an indoor nuisance species (Wilson and Taylor 1967), but is not a risk for structural damage. According to New Zealand records, the species is among the most commonly intercepted ants in that country (Ward et al. 2006). Sixty-nine percent of the interceptions were in freight from Fiji (> 92% from the Pacific Islands). Interceptions were mostly in fresh produce (69%) and cut flowers (8%). Pheidole fervens was also intercepted multiple times in air passengers’ luggage and shipping containers. The species could become more globally widespread in the future.
Diagnosis of worker among Antkey species. Worker castes bimorphic. Head shape ovoid (minor workers) or subrectangular with posterolateral lobes (majors), but never triangular. Antenna 12-segmented. Antennal club 3-segmented. Antennal insertions at least partly covered by frontal lobes; not surrounded by a raised sharp-edged ridge. Frontal lobes do not obscure face outline between mandible and eye; relatively close together so that the posteromedian portion of the clypeus, where it projects between the frontal lobes, is at most only slightly broader than one of the lobes. Posterolateral corners of head unarmed, without spines. Mandibles triangular. Mesosoma with erect hairs. Pronotal spines absent. Propodeum armed with spines or teeth. Slope of mesosoma steep. Waist 2-segmented. Petiole pedunculate with a distinct and upright node; lacking large subpetiolar process. Postpetiole attached to lower surface of gaster; not swollen; in dorsal view not distinctly broader than long or distinctly wider than petiole. Minor worker characters. Head ovoid; distinctly longer than wide; smooth and shining, lacking punctation. Antennal scrobe lacking. Antennal scapes extend beyond posterior margin of head. Posterior head margin weakly convex. Postpetiole not swollen relative to petiole. Lateral portions of petiole and postpetiole weakly sculptured to lacking sculpture. Color yellowish brown. Major worker characters. Head shape rectangular (HL > HW). Posterolateral lobes distinctly rugose. Antennal scrobe weakly to moderately impressed, but some depression capable of receiving antennal scapes clearly visible. Rugae above eye level moderately reticulated. Promesonotum in profile with two distinct convexities.
Diagnosis among introduced Pheidole. Color yellowish brown to dark brown. Major | HW 1.13–1.44, HL 1.13–1.56, SL 0.80–0.95, CI 92–100, SI 61–71 (n=15, Eguchi 2001a; 2008; Fischer and Fisher 2013). Head square to subquadrate (Fig. 7); rugoreticulate on posterolateral lobes and laterad of frontal carinae (Fig. 13a), but frons dominated by long, well-organized and parallel longitudinal rugae (Fig. 13b). Antennal scrobes indistinct to moderately impressed, but frontal carinae always forming a border capable of accepting the antennal scape (Fig. 13c). Frontal carinae relatively longer, extend 4/5 distance of head before terminating (Fig. 14). Promesonotum in profile with two convexities (Fig. 5), the large anterior dome in addition to a distinct mound or prominence on the posterior slope. Postpetiole not swollen relative to petiole (Fig. 3). Minor | HW 0.52–0.63, HL 0.66–0.73, SL 0.77–0.87, CI 79–88, SI 133–154 (n=16, Eguchi 2001a; 2008; Fischer and Fisher 2013). Head predominantly glossy (Fig. 36), lacking punctation or rugulae above eye level. Posterior head margin weakly convex to flat in full-face view (Fig. 45). Antennal scapes long (e.g. Fig. 39), but not surpassing the posterior head margin by more than 2x eye length. Promesonotum in profile with two convexities, the large anterior dome (Fig. 43a) in addition to a distinct prominence on the posterior slope (Fig. 43b). Promesonotal prominence relatively flat (Fig. 49a). Metanotal depression relatively deep (Fig. 49b). Petiole and postpetiole glossy to very weakly sculptured laterally (Fig. 48). Postpetiole not swollen relative to petiole (Fig. 3).
Pheidole indica
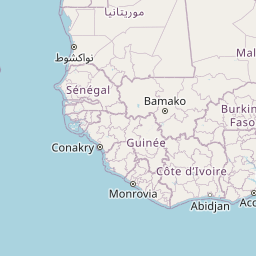
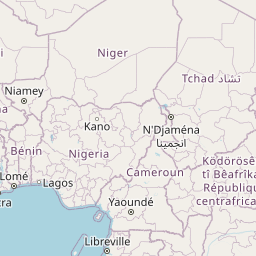

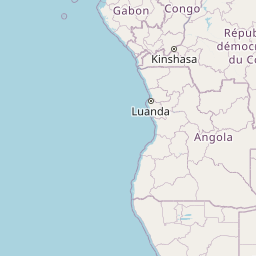

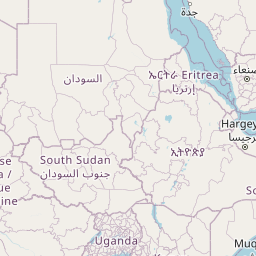
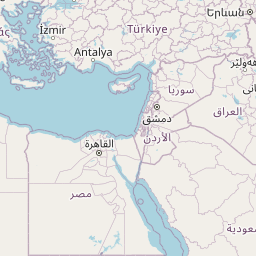

We consider Pheidole fervens as native to a broad expanse of the Indo-Malay region spanning from India east to the Philippines and south to the islands west of New Guinea. This is a broad and admittedly arbitrary boundary, but a more precise circumscription of the native range requires a population-level analysis outside the scope of the present study. In particular, it is difficult to ascertain the extent of its range into the Pacific Island region prior to the Anthropocene. The only known occurrence of P. fervens from New Guinea was a single record from the westernmost part of the island (Emery 1887b). East of New Guinea, however, the species is established on nearly all islands of the Pacific, including those which were uninhabited by any ant prior to human arrival. Although it is quite possible that P. fervens reached some of these islands without human assistance—especially those between Taiwan and mainland Japan—we treat these as introduced populations. And although established on Mauritius, the species is rarely encountered there and is currently known from only two localities (Fischer and Fisher 2013). The only record of introduction in North America is a California population that established nests in cracks of roads and along the sides of buildings in a two-block area of downtown Los Angeles (Martínez 1996). Pheidole fervens has been collected from greenhouses in the Netherlands (Boer and Vierbergen 2008), and is frequently intercepted by quarantine inspections (Ward et al. 2006).